生理学/脊髓对躯体运动的调节
| 医学电子书 >> 《生理学》 >> 神经系统 >> 神经系统对躯体运动的调节 >> 脊髓对躯体运动的调节 |
| 生理学 |
|
|
|
在脊髓的前角中,存在大量运动神经元(α和γ运动神经元),它们的轴突经前根离开脊髓后直达所支配的肌肉。α运动神经元的大小不等,胞体直径从几十到150μm;大α运动神经元支配快肌纤维,小α运动神经地支配慢纤维。α运动神经元接受来自皮肤、肌肉和关节等外周传入的信息,也接受从脑干到大脑皮层等主位中枢传的信息,产生一定的反射传出冲动。因此,α运动神经元是躯干骨骼肌运动反射的最后公路。
α运动神经元的轴突在离开脊髓走和肌肉时,其末梢在肌肉中分成许多小支,每一小支支配一根骨骼肌纤维。因此,在正常情况下,当这一神经元发生兴奋时,兴奋可传导到受它支配的许多肌纤维,引起其收缩。由一个α运动神经元及其支配的全部肌纤维所组成的功能单位,称为运动单位。运动单位的大小,决定于神经元轴突开梢分支数目的多少,一般是肌肉愈大,运动单位也愈大。例如,一个眼外肌运动神经元只支配6-12根肌纤维,而一个肢肌(如三角肌)的运动神经元所支配的肌纤维数目可达2000根。前者有利于肌肉进行精细的运动,后者有利于产生巨大的肌张力。同一个运动单位的肌纤维,可以和其他运动单位的肌纤维交叉分布,使其所占有的空间范围比该单位肌纤维截而的总和大10-30倍;因此,即使只有少数运动神经元活动,在肌肉中产生的张力也是均匀的。
γ运动神经元的胞体分散在α运动神经元之间,其胞体较α运动神经元为小。γ运动神经元的轴突也经前根离开脊髓,支配骨骼肌骨的梭内肌纤维。据观察,前根中神经纤维的三分来自γ运动神经元。γ运动神经元的兴奋性较高,常以较主频率持续放电。在安静和麻醉的运动中都观察到,即使α运动神经元无放电,一些γ运动神经元仍持续放电。γ运动神经元和α运动神经元一样,末梢也是释放乙酰胆碱作为递质的。在一般情况下,当α运动神经元活动增加时,γ运动神经元也相应增加,从而调节着肌梭对牵拉刺激的敏感性。
目录 |
(一)脊休克
在动物中将脊髓与延髓的联系切断,用以研究脊髓单独的功能;但为了保持动物的呼吸功能,常在颈脊髓第五节水平以下切断,以保留膈神经对膈肌呼吸的传出支配。这种脊髓与高位中枢离断的动物称为脊动物。在脊动物可以观察到脊髓的一些基本功能;但由于失去了高位中枢的调节,因而不能完全反映正常的脊髓功能。
与高位中枢离断的脊髓,在手术后暂时丧失反射活动的能力,进入无反应状态,这种现象称为脊休克(spinal shock)。脊休克的主要表现为:在横断面以下的脊髓所支配的骨骼肌紧张性减低甚至消失,血压下降,外周血管扩张,发汁反射不出现,坡肠和膀胱中粪尿积聚,说明动物躯体与内脏反射活动均减退以至消失。脊休克现象只发生在切断水平以下的部分。以后,一些以脊髓为中枢的反射活动可以逐渐恢复,恢复的迅速与否,与动物种类有密切关系;低等动物如蛙在脊髓离断后数分钟内反射即恢复,在犬则需几天,而在人类则需数周以至数月(人类由于外伤等原因也可出现脊休克)。显然,反射恢复的速度与不同动物脊髓反射依赖于高位中枢的程度有关。反射恢复过程中,首先是一引起比较简单、比较原始的反射先恢复,如屈肌反射、腱反射等;然后才是比较复杂的反射逐渐恢复,如对侧伸肌反射、搔爬反射等。反射恢复后的动物,血压也逐渐上升到一定水平,动物可具有一定的排粪与排快活反射,说明内脏反射活动也能部分地恢复。反射恢复后,有些反射反应比正常时加强并广泛扩散,例如屈肌反射、发汗反射等。
脊休克的产生并不由于切断损伤的刺激性影响引起的,因为反射恢复后进行第二次脊髓切断损务并不能使脊休克重现。所以,疹休克的产生原因是由于离断的脊髓突然失去了高位中枢的调节,这里主要指大脑皮层、前庭核和脑干网状结构的下行纤维对脊髓的易化作用。
脊休克的产生与恢复,说明脊髓可以完成某些简单的反射活动,但正常时它们是在高位中枢调节下进行活动的。高位中枢对脊髓反射既有易化作用的一面,也有抑制作用的一面。例如,切断脊髓后伸肌反射往往减弱,说明高位中枢对脊髓伸肌反射中枢有易化作用;而发汗反射加强,又说明高位中枢对脊髓发汗反射中枢有抑制作用。
脊动物由于脊髓离断,脊髓内上行与下行的神经束秀难重新接通,造成感觉传入冲动不能上达大脑皮层,而大脑皮层的传出冲动也不能下达脊髓,以致在离断水平以上就失去了知觉和所谓随意动作。中枢的神经轴突与外周的神经轴突都有再生能力,但由于脊髓离断部位有大量胶质细胞浸润并逐步形成瘢痕,使轴突再生受阻。
脊髓离断后屈肌反射比正常时加强,而伸肌反射往往减弱,以致屈肌反射常占优势,这不利于瘫痪肢体支持体重。因此,在低位脊髓横贯性损伤的病人,通过站立姿势的积极锻炼以发展伸肌反射是很重要的;这种锻炼使下肢伸肌具有足够的紧张性以保持伸直,以使不依靠拐杖站立或行走。同时,通过锻炼充分发挥末瘫痪肌肉的功能,例如背阔肌等由脊髓离断水平以上的神经所支配,而却随着于骨盆,这样就有可能使病人在借拐杖行走时摆动骨盆。
(二)屈肌反射与对侧伸肌反射
在脊动物的皮肤接受伤害性刺激时,受刺激一侧的肢体出现屈曲的反应,关节的屈肌收缩而伸肌驰缓,称为屈肌反射。屈肌反射具有保持性意义。屈肌反射的强度也刺激强度有关,例如足部的较弱刺激只引致踝关节屈曲,刺激强度加大,则膝关节及髋关节也可发生屈曲。如刺激强度更大,则可以同侧肢体发生屈肌反向的基础上出现对侧肢体伸直的反射活动,称为对侧伸肌反射。对侧伸肌反射是姿势反射的之一,具有维持姿势的生理意义,动物一侧肢体屈曲,对侧肢体伸直以支持体重。屈肌反射是一种多突触反射,其反射弧传出部分可通向许多关节的肌肉。
在人类由于锥体束或大脑皮层运动区的功能发生障碍,脊髓失去了运动区的调节,可出现一处特殊的反射。例如,以钝物划足跖外侧时,出现大趾背屈,其他四趾向外展开如扇形的反射,称为巴彬斯基征(Babinski’s sign)阳性。从生理学角度来看,这一反射属于屈肌反射,因为当刺激加强时还可伴有踝、膝、髋关节的屈曲。平时脊髓在大脑皮层的调节下,这一原始的屈肌反射被抑制而不表现出来。在婴儿的锥体束未发育完全以前,以及成人深睡或麻醉状态下,也可以出现巴彬斯基征阳性。
(三)牵张反射
有神经支配的骨骼肌,如受到外力牵拉使其伸长时,能产生反射效应,引起受牵扯的同一肌肉收缩,此称为牵张反射(stretch reflex)。牵张反射有两种类型,一种为腱反射,也称位相性牵张反射;另一种为肌紧张,也称紧张性牵张反射。
腱反射是指快速牵拉肌腱时发生的牵张反射。例如,叩击膝关节下的股四头肌腱使之受到牵扯,则股四头肌即发生一次收缩,这称为膝反射;叩击跟腱使之受到牵扯,则小腿腓肠肌即发生一次收缩,这称为跟腱反射。这些腱反射的感受器为肌梭,传入神经纤维的直径较粗(12-20μm)、传导速度较快(90m/s以上),效应器为同一肌肉的肌纤维;反射反应的潜伏期很短,据测算兴奋通过中枢的传布时间公0.7ms左右,只够一次突触接替的中枢延搁时间。因此,腱反射为单突触反射,传入神经纤维经背根进入脊髓灰质后,直达前角与运动神经元发生突触联系。当叩击肌腱时,肌肉内的肌梭同时受到牵张,同时发动牵张反射。因此肌肉的收缩几乎是一次同步性收缩。腱反射主要发生于肌肉内收缩较快的快肌纤维成分。
肌紧张是指缓慢持续牵拉腱时发生的牵张反射,其表现为受牵拉拢肌肉能发生紧张性收缩,阻止被拉长。肌紧张是维持躯体姿势最基本的反射活动,是姿势反向的基础。例如,由于重力影响,支持体重的关节趋向于被重力所弯曲,关节弯曲必使伸肌肌腱受到持续牵拉,从而产生牵张反射引起该肌的收缩,对抗关节的屈曲,维持站立姿势。由于重力经常作用于关节,因此这种牵张反射也就持续着。肌紧张与腱反射的反射弧基本相似,感受器也是肌梭,但中枢的突触接替可能不止一个,即可能是多突触反射,其效应器主要是肌肉内收缩较慢的慢肌纤维成分。肌紧张的反射收缩力量并不大,只是抵抗肌肉被牵拉,因此不表现明显的动作。这可能是因为在同肌肉内的不同运动单位进行交替性的收缩而不是同步性收缩,所以肌紧张能持久维持而不易疲劳。
牵张反射主要是使受牵拉的肌肉发生收缩,但同一关节的协同肌也能发生兴奋,而同一关节的颉颃肌则受到抑制(交互抑制),但并不影响其它关节的肌肉,伸肌和屈肌都有牵张反射,但脊髓的牵张反射主要表现在伸肌。屈肌的牵张反射表现不明显,主要表现为它的颉颃肌(即伸肌)受到抑制。牵张反射(尤其是肌紧张)的主要生理意义在于维持这种站立姿势,因此伸肌比屈肌的牵张反射明显符合生理情况。
牵张反射的基本反射弧是简单的,但在整体内牵张反射是受高位中枢调节的,而且可以建立条件反射。腱反射的减弱或消失,常提示反射弧的传入、传出通路受脊髓反射中枢的损害或中断;而腱反射的亢进,则常提示高位中枢的病变I(例如锥体束综合症)。因此,临床上常用测定腱反射的方法来了解神经系统的功能状态。常用的腱反射见表10-5。
表10-5 常用的腱反射
| 名称 | 检查方法 | 中枢部位 | 效应 | |
| 肘反射 | 扣击肱二头肌肌腱 | 颈5-7 | 肘部屈曲 | |
| 膝反射 | 扣击髌韧带 | 腰2-4 | 小腿伸直 | |
| 跟腱反射 | 扣击跟腱 | 腰5-骶2 | 脚向足底方向屈曲 | |
肌梭和腱器官
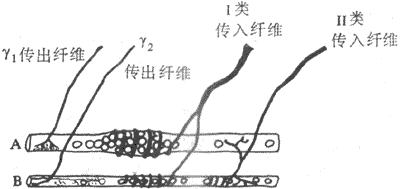
肌梭是一种感受肌肉长度变化或感受牵拉刺激的特殊的梭形感受装置,长几个毫米,外层为一结谛组织囊(图10-30)。肌梭囊内一般含有6-12根肌纤维,称为梭内肌纤维;而囊外的一般肌纤维就称为梭外肌纤维。整个肌梭现阶附着在梭外肌纤维上,并与其平等排列呈并联关系。梭内肌纤维的收缩成分位于纤维的两端,而感受装置位于其中间部,两者呈串联关系。因此,当梭外肌纤维收缩时,感受装置所受的牵拉刺激将减少;而当梭内肌纤维收缩时,则感受装置对牵拉刺激的敏感度增高。肌梭的传入神经支配有两类。I类传入纤维直径较粗(12-20μm),Ⅱ类传入纤维直径较细(4-12μm)。中枢有运动传出支配梭外肌和梭内肌纤维,前者称为α传出纤维(直径12-20μm),后者称为γ传出纤维(直径2-6μm)。当γ传出纤维活动加强时,梭内肌纤维收缩,可提高肌梭内感受装置的敏感性,因此γ传出纤维的活动对调节牵张反射具有重要作用。进一步研究指出,梭内肌纤维分两类:一类其细胞核集中于中央部称为核袋纤维(nuclear bag fiber),它接受γ1传出纤维支配,并对快速牵拉较敏感,其传入纤维主要属I类;另一类其细胞核分散于整个纤维称为核链纤维(nuclearchain fiber),它接受γ2传出纤维支配,并对缓慢持续牵拉较敏感,其传入纤维有I类和Ⅱ类(图10-31)。
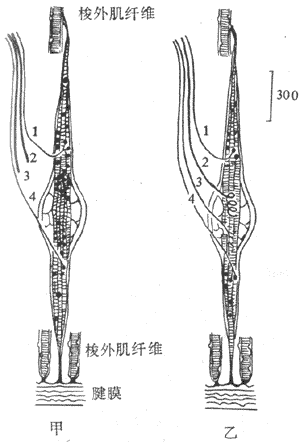
图10-31 两类梭内肌纤维示意图
A:核袋纤维 B:核链纤维
腱器官分布在肌腱胶原纤维之间的牵张感受装置,由较细的I类纤维(直径12μm)支配,末梢一般只有几个分支。腱器官与梭外肌纤维呈串联关系,其功能与肌梭功能不同,是感受骨肉张力变化的装置。当梭外肌纤维发生等长收缩时,腱器官的传入冲动发放频率不变,肌梭的传入冲动频率减少;当肌肉受到被动牵拉时,腱器官和肌梭的传入冲动发放频率均增加。因此,腱器官是一种张力感受器,而肌梭是一种长度感受器。此外,腱器官的传入冲动对同一肌肉的α运动神经元起牵拉抑制作用,而肌梭的传入冲动对同一肌肉的α运动神经元起兴奋作用。一般认为,当肌肉受到牵拉时,首先兴奋肌梭的感受装置发动牵张反射,引致受牵拉的肌肉收缩以对抗牵拉;当牵拉力量进一步加大时,则可兴奋腱器官使牵张反射受抑制,以避免被牵拉的肌肉受到损伤。
(四)节间反射
脊动物在反射恢复的后期,可出现复杂的节间反射。例如,刺激动物腰背皮肤,可引致后肢发生一系列节奏性搔爬动作,称为搔爬反射。搔爬反向依靠脊髓上下节段的协同活动,所以是节间反射的一种表现。
| 关于“生理学/脊髓对躯体运动的调节”的留言: | |
|
目前暂无留言 | |
| 添加留言 | |