生物化学与分子生物学/蛋白多糖
| 医学电子书 >> 《生物化学与分子生物学》 >> 结缔组织生化 >> 蛋白多糖 |
| 生物化学与分子生物学 |
|
|
结缔组织基质中蛋白质与多糖以共价和非共价键相连构成多种巨大分子称为蛋白多糖(proteoglycans)或粘蛋白(mucoproteins)。其分子组成以多糖链为主,蛋白质部分所占比例较小。往往一条多糖链上联结多条多肽链,分子量可达数百万以上。
目录 |
一、化学结构
蛋白多糖中的多糖链为杂多糖,因其组成成分中均含氨基已糖,所以称为氨基多糖或糖胺聚糖(glycosaminoglycans)。人体组缔组织中常见的氨基多糖包括透明质酸(hyaluronicacid)、硫酸软骨素(chondroitin sulfate)、硫酸角质素(keratan sulfate)和肝素(heparin)等。
表13-2 人体中常见的糖胺多糖
| 名称 | 基本结构单位 | 主要存在部位 |
| 透明质酸 |
|
关节液、软骨、结缔组织基质、皮肤、脐带、玻璃体液 |
| 4-硫酸软骨素(硫酸软骨素A) |
|
骨、软骨、角膜、皮肤、血管 |
| 6-硫酸软骨素(硫酸软骨素C) |
|
软骨、肌腱、脐带、椎间盘 |
| 硫酸皮肤素(硫酸软骨素B) |
|
皮肤、韧带、动脉、心瓣膜 |
| 硫酸角质素 |
|
角膜、软骨、髓核 |
| 肝素 |
|
肺、皮肤、肝、肠等肥大细胞及嗜碱性白细胞内 |
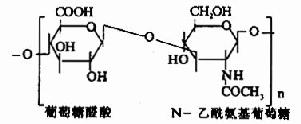
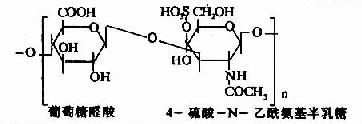
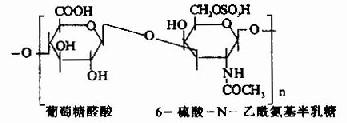
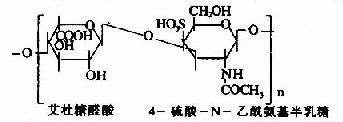
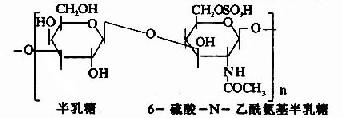
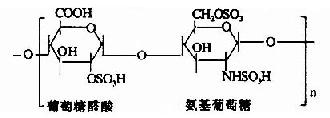
其中硫酸软骨素B又称为硫酸皮肤素(dermatan sulfate)。各种糖胺多糖的结构单元为二糖单位,均含有乙酰氨基已糖和糖醛酸(硫酸角质素例外,不含糖醛酸而代之以半乳糖)。除透明质酸外都含硫酸。因而糖胺多糖为酸性,加之分子大具有粘性,故又称为酸性粘多糖(acidmucopolysaccharide,AMPS)。
经电子显微镜图形及实验推测,蛋白多糖为“瓶刷状”分子结构(图13-1),其蛋白多糖亚单位(“刷毛”)非共价附着于透明质酸主链上(backbone),其间相隔200-300A。
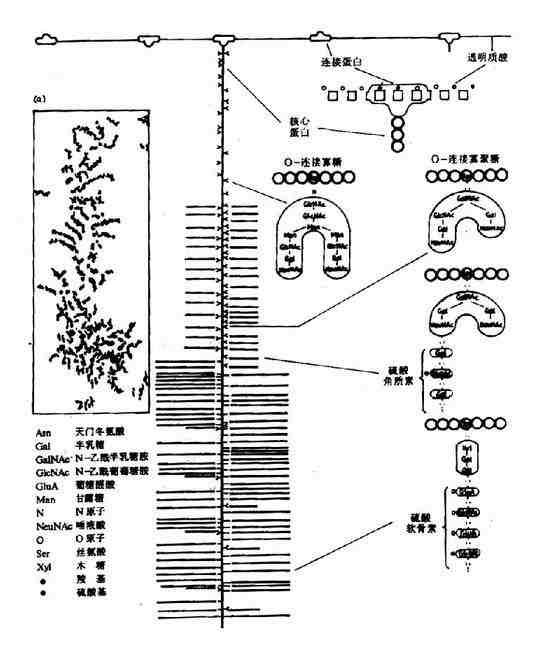
图13-1 蛋白多糖的分子结构
蛋白多糖亚单位由一个核心蛋白(coreprotein)和共价连接其上的糖胺多糖组成,后者主要为硫酸角质素和硫酸软骨素。人体中有多种不同的核心蛋白,分子量达200~300kD,是所有组织细胞中分泌的最大的一种多肽。核心蛋白高度伸展N-末端,形成一球状区,约60-70kD,非共价连接于透明质酸链上,另一种约40-60kD的连接蛋白(link protein)参与稳定球状区与透明质酸链的非共价连接。核心蛋白及多糖组成的亚单位可分为三区:
(1)N-端区:包括球状连接区,含有较少的寡糖链。
(2)富含寡糖区:为硫酸角质素寡糖链的主要附着区。寡糖链共价连接于核心蛋白分子中丝氨酸和苏氨酸残基侧链氧原子上。
(3)C-末端区:富含硫酸软骨素。通过半乳糖半乳糖木糖三糖连接于核心蛋白的丝氨酸残基。
蛋白多糖的透明质酸主链长度为4000~40,000,可附着上百个核心蛋白,每条核心蛋白可结合50条硫酸角质素链和100条硫酸软骨素链,由此可见蛋白多糖分子巨大,分子量可高达数千万。
二、蛋白多糖的生理功能
蛋白多糖分子大,具高度亲水性,对保持结缔组织水分及与组织间物质交换均有重要作用。例如软骨组织中胶原纤维排列成网格状,网格间隙中填充蛋白多糖,因其有高度亲水性,吸附大量水份在其中,当软骨受压时,水分可被挤压出去,而减压后又可重吸进来。关节软骨无血管供应,其营养物质的交换主要靠运动产生压力变化使液体流动。由此可以解释长期不运动导致关节软骨的萎缩。
蛋白多糖的糖链上含有较多的酸性基团,对于细胞外液中Ca++、Mg++、K+、Na+等阳离子有较大的亲和力,因此能调节这些阳离子在组织中的分布。
蛋白多糖分子巨大,有较大的粘滞性,附着于组织表面,能缓冲组织之间的机械磨擦,因而具有润滑、保护作用。例如关节液中的蛋白多糖有润滑关节面的作用。类风湿关节炎患者,关节液中蛋白多糖减少,失去原有润滑作用,因而关节面易被破坏。
蛋白多糖与创伤的愈合亦有密切关系。皮肤创伤后的肉芽形成过程中,通常先有糖胺多糖的增生,进而促进胶原纤维的合成,其机理尚不清楚。近年来发现蛋白多糖可调节某些蛋白生长因子的作用。例如碱性成纤维细胞生长因子(basic fibrotlast growth factor,bFGF)结合于蛋白多糖的肝素或硫酸角质素链上,而且要以bFGF糖胺多糖复合物形式与细胞表面受体结合。bFGF与肝素或硫酸角质素结合能保护bFGF免被降解。当核蛋白水解或肝素或硫酸角质素链部分降解时可释放出活性bFGF糖胺多糖复合物,刺激细胞增殖及分化。其它一些生长因子也有类似作用机制,很明显,广泛和大量存在的蛋白多糖可局限这些生长因子在其分泌细胞短距离内作用。这对于组织的特有结构具有重要意义。
三、蛋白多糖的生物合成
蛋白多糖的合成首先按蛋白质生物合成的原理,在核糖体上合成多肽,并分泌入内质网中,在内质网中修饰过程中,由相应的转移酶催化活性单糖转移到氨基酸的侧链上,合成胺基多糖。但糖链的延伸和加工修饰在高尔基体进行。所以说多肽的合成受专一基因控制,而氨基多糖的合成主要由酶的分隔定位和对酶特异性所决定。
参与胺基多糖合成的各种单糖及其衍生物需先活化成活性单糖,即与二磷酸尿苷(UDP)结合,而各种单糖及其衍生物均可由葡萄糖转变而来,如下图13-2所示:
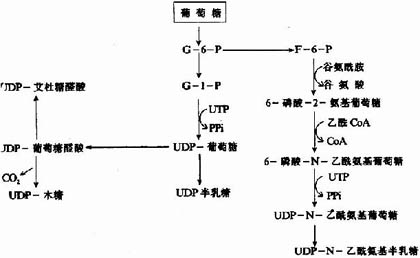
图13-2 各种单糖及其衍生物的来源
糖胺多糖合成的起始步骤是在木糖转移酶(Xylose transfevase)的催化下,将一分子木糖基连接到核心蛋白多肽链的丝氨酸残基上,形成O-糖苷键(O-glycosidicbond)。再由半乳糖转移酶(galactose transforase)催化依次转移两分子半乳糖,构成“木棸霔半”三糖连接区。然后再由高度特异的糖基转移酶作用逐渐按顺序延长,糖链合成后再进一步修饰。由差向异构酶催化将葡萄糖醛酸转变为艾杜糖醛酸,而硫酸基则是由活性硫酸根(PAPS)提供,由硫酸转移酶
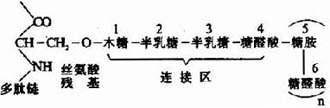
图13-3 糖胺多糖链与多肽链的连接
催化加到新合成的多糖链的氨基或羟基上。(图13-4为硫酸软骨素A合成过程示意图),当Vit A缺乏时硫酸转移酶活性下降,糖胺多糖合成受限,可引起组织生长、发育的障碍。
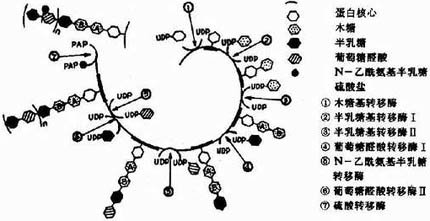
图13-4 硫酸软骨素A的合成示意
四、蛋白多糖的分解代谢
结缔组织基质中的蛋白多糖主要受组织蛋白酶D等的作用,部分肽链水解产生的带多糖链的小片段可被细胞吞噬,进而在溶酶体中逐步水解成各种单糖及其衍生物。因此,溶酶体是糖胺多糖分解的主要场所。
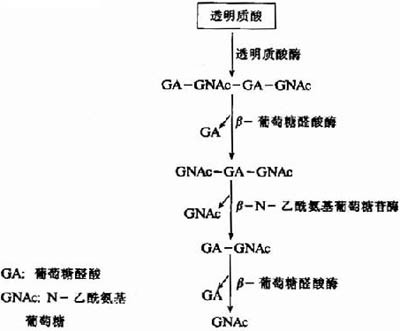
图13-5 透明质酸的水解
溶酶体中分解糖胺多糖的酶包括内切糖苷酶、外切糖苷酶及硫酸酯酶等。如图13-5所示透明质酸的水解过程:首先透明质酸酶(hyaluronidase)为一种内切酶,能水解透明质酸、硫酸软骨素A和C中的β-N-乙酰氨基已糖糖苷键,产生主要为四糖或六糖的寡糖。随后再由β-葡萄糖醛酸酶及β-N-乙酰氨基葡萄糖苷酶等外切酶进一步水解,成为单糖及其衍生物。
大多数糖胺多糖都含有硫酸基团。溶酶体中也存在多种硫酸酯酶,能水解各种硫酸酯键。有些糖苷酶有很强的底物特异性、不能水解带硫酸基的寡糖链,因此,硫酸酯酶的先天性缺乏会引起糖胺多糖分解障碍,导致粘多糖症(mucoplysaccharidoses)。
参看
| 关于“生物化学与分子生物学/蛋白多糖”的留言: | |
|
目前暂无留言 | |
| 添加留言 | |