营养学/碳水化物的生化代谢
| 医学电子书 >> 《临床营养学》 >> 碳水化物 >> 碳水化物的生化代谢 |
| 临床营养学 |
|
|
|
4.2.1 消化吸收
碳水化物要消化或单糖才吸收。消化的过程就是水解的过程。麦芽糖、乳糖、蔗糖、麦芽低聚糖都能消化。人能消化的多糖仅淀粉一种,糖原在制成食品时已不存在了。
消化从口腔开始,口腔里有唾液淀粉酶能水解交替α1→4糖甘键,但不能水解α1→6糖苷键和相邻的α1→6糖苷键。消化产物是糊精、麦芽低聚糖和麦芽糖。
胃里没有消化淀粉的酶。唾液淀粉酶的最适PH是6.6~6.8,在食糜没有被胃酸中和以前,能持续作用一段时间,使淀粉和低聚糖能再消化一部分。
小肠内有胰液的α-淀粉酶,其作用和唾液淀粉酶相同,把直链淀粉消化成麦芽糖和麦芽三糖,支链淀粉消化成麦芽糖、麦芽三糖及由4~9个葡萄糖分子组成的而有α1→6苷键的麦芽低聚糖。
肠粘膜上皮细胞中有吸收细胞,每一细胞约有3000条微绒毛,微绒毛间的空间的有效半径约0.4nm。只有上述消化产物能够通过,与微绒毛膜上的酶反应。膜上的酶有四种:①α1→4糖苷酶,把葡萄糖分子自上述产物一个个地切下来;②异麦芽糖酶,水解麦芽低聚糖的α1→6糖苷键;③蔗糖酶,消化蔗糖;④β-半糖苷酶,消化乳糖。
所以消化分两步进行:①肠腔内的消化,产物是双糖和麦芽低聚糖;②微绒毛膜上的消化,产物是单糖。
微绒毛双脂质层的(详后)。四种酶嵌在双脂质层内,活性位伸在膜外。在其近处,还有全部嵌在膜内的运输单糖的蛋白质,这样,消化的最终产物立刻可以被运输蛋白所结合。
运输蛋白在结合葡萄糖以前,先结合肠腔内的Na+排入肠腔,肠腔中Na+都带入细胞内,释放到胞浆中。Na+排入肠腔,肠腔中的Na+浓度比细胞内高,自低浓度排到高浓度要消耗能量,所需能量由ATP供应。糖进入细胞后,约有15%流回肠腔,25%扩散入血,60%与靠近基膜一端的质膜上的另一载体蛋结合而离开细胞。这一结合不需Na+,而且运输葡萄糖的速度比葡萄糖从肠腔进入吸收细胞的速度快,所以葡萄糖不会在吸收细胞中蓄积,从而提高了吸收效率。当食糜到达空肠下部时,95%的碳水化物都被吸收了。
4.2.2 中间代谢概述
小肠吸收的碳水化物主要是葡萄糖、果糖、半乳糖三种,经门静脉送到肝脏。葡萄糖进入肝细胞后与磷酸反应生成葡萄糖-6-磷酸,这样细胞内的葡萄糖浓度可维持在低水平,使葡萄糖不断进入肝细胞。在吸收的葡萄糖中,60%以上在肝内代谢,其余入大循环。果糖和半乳糖在肝中转变为葡萄糖。
葡萄糖在肝内经分解代谢提供机体所需要的能量,多余的合成糖原保留在肝内,再有多余就转变成脂肪运送到脂肪组织储存起来。
葡萄糖的分解代谢分三个阶段:
(1)酵解葡萄糖分解的第一阶段是生成丙酮酸。这一系列反应和碳水化物在体外经酵母的作用生成丙酮酸相同,所以称为酵解。所不同的是发酵时丙酮酸进一步生成洒精;在体内,有氧时丙酮酸氧化成二氧化碳和水,无氧时还原成乳酸。
酵解分步进行(图4-11)。为了要把葡萄糖的6个碳原子一分为二,先使它两端磷酸化,才能断列成两个接近相等的部分,然后脱去磷酸成丙酮酸。
上述反应都在细胞浆中进行。每一步都有特定的酶参加。第①、③两步消耗2分子ATP,说明葡萄糖分解代谢开始时要用能量来推动。⑥、⑨两步各产生2分子ATP。从葡萄到丙酮酸,净生2分子ATP,酵解产生的能量不多。
细胞浆中NAD+和NADH的总量基本恒定。如所有的NAD+都变成NADH,酵解就得停止。要使葡萄继续不断氧化成酮酸,必需把NADH的氢转移到另外的受体上去。有氧时氢经呼吸链氧化成水。无氧时氢交给丙酮酸使成乳酸⑩。乳酸过多时离开细胞入血。所以严重缺氧者可发生代谢性酸中毒。
(2)丙酮氧化成乙酰酶a 有氧时,丙酮酸进入线粒体。在丙酮酸脱氢酶系的帮助下,氧化脱羟成乙酰辅酸脱氢酶系的帮助下,氧化脱羟成乙酰辅酶A,同时产生1molATP和1molNADH。维生素B1B2、烟酰胺、泛酸参加。反应是不可逆的。
(3)三羧酸循环乙酰辅酶A上的两个来自葡萄糖的碳原子,经三羟酸循环生成2molCO2,完成葡萄糖的氧化。反应在线粒体中进行。这一过程分步(图4-12)。
三羧酸循环中产生3molNADH,1molFADH2,1molGTP。
NADH把氢通过FAD和细胞色素交给氧,同时放出能量。每molNADH通过这一系列反应所放出的能量是220kJ(52.6kCal)。这些能量以ATP的形式贮存起来。每molATP能产生30.5Kj(7.3kCal),理论上每molNADHRNUD氧化产生的能量可用来合成7molATP。这是指在标准状况(25℃,1个大气压,作用物和产物浓度都是1M/LPH7)下说的。在模拟体内的条件时,实验证明只能合成3molATP。如用反应式表示:
NADH+H++1/2O2
![]() NAD++H2O+能量
NAD++H2O+能量
3ADP+3H3PO4+能量
![]() 3ATP+3H2o
3ATP+3H2o
前面一个是氧化反应,后面一个是磷酸化,二者偶合在一起,称为氧化磷酸化。
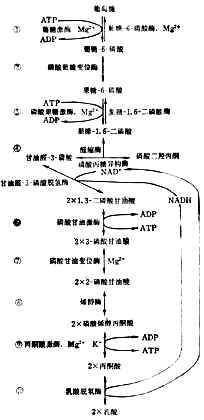
图4-11 糖的酵解
实验也证明FADH2氧化时产生2个ATP,GTP可生成1molATP。根据这些数值,葡萄糖完全氧化时,机体获得的能量可归纳如下:
①酵解:葡萄糖→2丙酮酸+8ATp
②丙酮酸氧化成乙酰辅酶A:
2丙酮酸→2乙酰辅酶A+2CO2+6ATp
③三羧酸循环:
2乙酰辅酶A→4CO2+24ATp
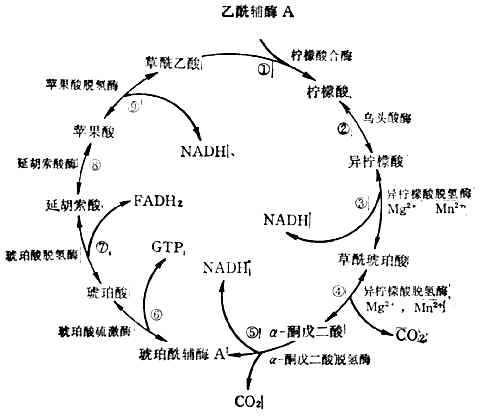
图4-12三羧酸循环
即每mol葡萄糖在体内氧化成6molCO2时可产生38molATP,机体可以利用的能量是38×30或1140kJ[(38×7.3)或277kCal]。每mol葡萄糖氧化成6molCO2和水时,自由能的改变是2870KJ(686kCal)。所以,机械效率=1140/2870×10=40%。
这是利用标准状况的数字计算的,在体内高一些,约60%。
从上面归纳的数字也可看出碳水化物提供的能量,在氧化的三个阶段中,以三羧酸循环最多。脂肪和蛋白质最后也通过三羧酸循环氧化。食物向机体提供的能量,90%来自三羧酸循环。
葡萄糖代谢另外还有一种途径,不通过果糖二磷酸,叫做已糖一磷酸通路。由于产生核榶,又叫做戊糖-磷酸通路。
(4)已糖-磷酸通路这一通路目的是提供合成核酸所需的核糖,以及合成脂肪酸、胆固醇等所需的能量供应者NADPH(还原型辅酶Ⅱ)(图4-13)。肝脏中约有30%的葡萄糖经过这一通过代谢。红细胞中这一通路也很活跃。它的失调可导致溶血性贫血。反应都在细胞浆中进行。
核糖-5-磷酸既可用于核酸的生物合成,又可与5-磷酸木酮糖反应生成甘油醛3-磷酸和果糖-6-磷酸。究竟走哪一条路,要看细胞本身对NADPH和核糖的需要量而定。许多细胞NADPH要得多,过剩的戊糖可经甘油醛-3-磷酸和果糖-6-磷酸酵解,或由果糖-6-磷酸转变成葡萄糖-6-磷酸而重走这一通路。
这一通路需维生素B1参加。要使此路畅通,膳食中需有足量的维生素B1。5-磷酸核酮糖转变为葡萄糖-6-磷酸的反应,可用于评价维生素B1参加。要使此路畅通,膳食中需有足量的维生素B1的营养状况。
(5)糖原葡萄糖到各个细胞发挥作用以后,多余的可储存起来。避免细胞内渗透压的升高,细胞把葡萄糖合成大分子的糖原。需要能量时糖原再分解葡萄糖。如肝糖原产生的葡萄糖,用以维持血糖的恒定。肌糖原产生的葡萄糖,直接用于供给能量。
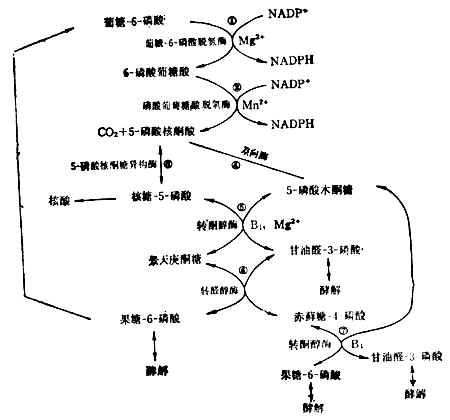
图4-13 已糖-磷酸通路
4.2.3 三大营养代谢的关系
人习惯于三餐,饱腹和空腹交替产生。所以要具备能处理碳水货物增多和缺少时保证能量供应的两套本领。
餐后血糖升沿高,胰岛素分泌增加,胰高血糖素分泌减少。使更多的葡萄糖进入肝脏、肌肉和脂肪组织;增强酶活力,加速葡萄的氧化和肝糖原、肌糖原的合成。超过糖原的储存量后,肝脏可把葡萄糖经磷酸二羟丙酮还原成甘油-3-磷酸,与乙酰辅酶A合成的脂肪酸,利用NADPH提供的能量,生成脂肪。再和蛋白质结合成极低密度脂蛋白入血,运送到脂肪组织储存。过多的碳水化物也能合成某些非必需氨基酸。
餐后约4h,不能利用食物中的葡萄糖。胰高血糖素分泌增加,胰岛素减少,转为利用肝糖原以供给血糖。肝糖原储存量不多,成人能动用的不到70g,不够一个晚上的消耗。所以,从餐后8h开始,还得利用脂肪以提供能量。甘油可变成糖,但在脂肪中所占重量不多。脂肪酸只能供能,不能转变为葡萄糖。因此,血糖的来源不得不转向氨基酸了。一般说来,除亮氨酸和赖氨酸外,其他氨基酸都是能成糖的,但从能量的观点看,只有丙氨酸和谷氨酸可作葡萄糖的来源。此时血糖的90%来自丙氨酸,10%来自谷氨酸。
肝脏能把肌肉中送来的丙氨酸作用成丙酮酸而生成葡萄糖。空腹时,肌肉中进入血液的氨基酸增加,其总量的30~40%是丙氨酸。但肌肉蛋白质的丙氨酸含量不到10%。另外的丙氨酸从哪里来的?葡萄糖经酵解可生丙酮酸,再经转氨作用丙氨酸。但这样葡萄糖的量并没有增加。增加的丙氨酸实际来自其他氨其酸。
肾脏把谷氨酸去氨后得α酮戊二酸,再和运来的成糖氨基酸如丙酸起转氨作用而得丙酮酸,最后得葡萄糖。α-酮戊二酸也能经三羟酸循环形成苹果酸,最后成葡萄糖。
| 关于“营养学/碳水化物的生化代谢”的留言: | |
|
目前暂无留言 | |
| 添加留言 | |