生理学/视杆细胞的感光换能机制
| 医学电子书 >> 《生理学》 >> 感觉器官 >> 视觉器官 >> 视杆细胞的感光换能机制 |
| 生理学 |
|
|
|
从上世纪末开始,有人就从视网膜中提取出了一定纯度的感光色素即视紫红质,它在暗处呈红色;实验中还可以证明,提取出来的这种感光色素对不同波长光线的吸收光谱,基本上和晚光觉对光谱不同部分的敏感性曲线相一致(图9-7)。这一事实十分重要,因为既然光线对某种感光色素的光化学作用的强度正好与这些光线所引起的视觉的强度相一致,那就是提示前者可能是后者的基础。
(一)视紫红质的光化学反应及其代谢
视紫红质的分子量约为27-28kd,是一种与结合蛋白质,由一分子称为视蛋白(opsin)的蛋白质和一分子称为视黄醛(retnal)的生色基团所组成。视蛋白的肽链序列已搞清,它的肽链中有7段穿越所在膜结构、主要由疏水性氨基酸组成的α-螺旋区段,同一般的细胞膜受体具有类似的结构。视黄醛由维生素A变来,后者是一种不饱和醇,在体内一种酶的作用下可氧化成视黄醛。提纯的视紫红质在溶液中对500nm波长的光线吸收能力最强,这与人眼在弱光条件下对光就业上蓝绿光区域(相当于500nm波长附近)感觉最明亮(不是感到了蓝绿色)的事实相一致(图9-7),说明人在暗视觉与视杆细胞中所含视紫红质的光化学反应有直接的关系。
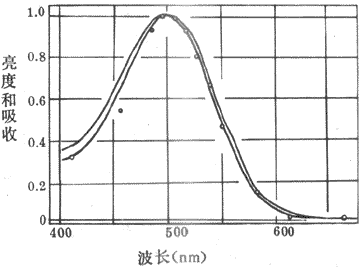
图9-7弱光条件下人眼所感到的光谱亮度曲线和实验条件下
视紫红质对光谱不同部分的吸收曲线视觉中最明亮的区域和视紫红制裁
吸收能力最强的部分都在500nm的波长附近
视紫红质在光照时迅速分解为视蛋白和视黄醛,这是一个阶段的反应。目前认为,分解的出现首先是由于视黄醛分子在光照时发生了分子构象的改变,即它在视紫红质分子中本来呈11-顺型(一种较为弯曲的构象),但在光照时变为全反型(一种较为直的分子构象)。视黄醛分子构象的这种改变,将导致视蛋白分子构象也发生改变,经过较复杂的信号传递系统的活动,诱发视杆细胞出现感受器电位。据计算,一个光量子被视紫红质吸收,就足以使视黄醛分子结构发生改变,导致视紫红质最后分解为视蛋白和视黄醛。视紫红质分解的某些阶段伴有能量的释放,但这看来不是诱发感受器电位的直接原因。
在亮处分解的视紫红质,在暗处又可重新合成,亦即它是一个可逆反应,其反应的平衡点决定于光照的强度。视紫红质再合成的第一步,是全反型的视黄醛变为11-顺型的视黄醛,很快再同视蛋白结合。此外,贮存在视网膜的色素细胞层中的维生素A也是全反型的,它们也可在耗能的情况下变成11-顺型的,进入视杆细胞,然后再氧化成11-顺型的视黄醛,参与视紫红质的合成补充;但这个过程进行的速度较慢,不是促进视紫红制裁再合成的即时因素。人在暗处视物时,实际是既有视紫红质的分解,又有它的合成,这是人在暗光处能不断视物的基础;光线愈暗,全盛过程愈超过分解过程,视网膜中处于合成状态的视紫红质数量也愈高,这也使视网膜对弱光愈敏感;相反,人在亮光处时,视紫红质的分解增强,合成过程甚弱,这就使视网膜中有较多的视紫红质处于分解状态,使视杆细胞几乎失去了感受光刺激的能力;事实上,人的视觉在亮光处是靠另一种对光刺激较不敏感的感光系统即视锥来完成的,后一系统在弱光时不足以被刺激,而在强光系统下视杆细胞中的视紫红质较多地处于分解状态时,视锥系统就代之而成为强光刺激的感受系统。在视紫红质和再合成的过程中,有一部分视黄醛被消耗,这最终要靠由食物进入血液循环(相当部分贮存于肝)中的维生素A来补充。长期摄入维生素A不足,将会影响人在暗光处的视力,引起夜盲症。
(二)视杆细胞外段的超威结构和感受器电位的产生
感光细胞的外段是进行光-电转换的关键部位。视杆细胞外段具有特殊的超威结构,如图9-8所示。在外段部分,膜内的细胞浆甚少,绝大部分为一些整齐的重叠成层的圆盘状结构所占据,这圆盘称为视盘。每一个视盘是一个扁平的囊状物,囊膜的结构和细胞膜类似,具有一般的脂质双分子层结构,但其中镶嵌着的蛋白质绝大部分是视紫红质,亦即视杆细胞所含的视紫红质实际上几乎全部集中在视盘膜中。视盘的数目在不同动物的视杆细胞中相差很大,人的每个视杆细胞外段中它们的数目近千;每一个视盘所含的视紫红质分子约有100万个。这样的结构显然有利于使进入视网膜的光量子有更大的机会在外段中碰到视紫红质分子。
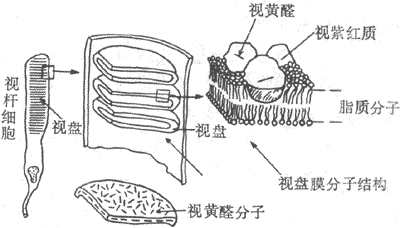
图9-8 视杆细胞外段的超微结构示意图
有人用细胞内微电极技术,研究了视杆细胞外段内外的电位差在光照前后的变化,结果发现在视网膜未经照射时,视杆细胞的静息电位只有-30?/FONT>-40mV,比一般细胞小得多。经分析表明,这是由于外段膜在无光照时,就有相当数量的Na+通道处于开放状态并有持续的Na+内流所造成,而内段膜有Na+泵的连续活动将Na+移出膜外,这样就维持了膜内外的Na+平衡。当视网膜受到光照时,可看到外段膜两侧电位短暂地向超极化的方向变化,由此可见,外段膜同一般的细胞膜不一致,它是在暗处或无光照时处于去极化状态,而在受到光刺激时,跨膜电痊反而向超极化方向变化,因此视杆细胞的感受器电位(视锥细胞也一样),表现为一种超极化型的慢电位,这在所有被研究过的发生器或感受器电位中是特殊的,它们一般都表现为膜的暂时去极化。
光子的吸收引起外段膜出现超极化电反应的机制已基本搞清,这就是光量子被作为受体的视紫红质吸收后引起视蛋白分子的变构,又激海参了视盘膜中一种称为传递蛋白(transducin)Ct的中介物,后者在结构上属于G-蛋白家庭的一员,它激活的结果是进而激活附近的磷酸二酯酶,于是使外段部分胞浆中的cGMP大量分解,而胞浆中cGMP的分解,就使未受光刺激时结合于外段膜的cGMP由也膜解离而被分解,而cGMP在膜上的存在正是这膜中存在的化学门控式Na+通道开放的条件,膜上cGMP减少,Na+通道开放减少,于是光照的结果出现了我们记录到的超极化型感受器电位。据估计,一个视紫红质被激活时,可使约500个传递蛋白被激活;虽然传递蛋白激活磷酸二酯酶是1对1的,但一个激活了的磷酸二酯酶在一秒钟内大约可使4千多个cGMP分子降解。由于酶系统的这种生物放大作用,就可以说明1个光量子的作用何以能在外段膜上引起大量化学门控式Na+通道的关闭,引起一个足以为人的视觉系统所感知的超极化型电变化。
视杆细胞外段和整个视杆细胞都没有产生动作电位的能力,由光刺激在外段膜上引起的感受器电位只能以电紧张性的扩布到达它的终足部分,影响终点(相当于轴突末稍)外的递质释放。
| 关于“生理学/视杆细胞的感光换能机制”的留言: | |
|
目前暂无留言 | |
| 添加留言 | |