临床生物化学/载脂蛋白的基因结构及表型
| 医学电子书 >> 《临床生物化学》 >> 血浆蛋白及其代谢紊乱 >> 载脂蛋白 >> 载脂蛋白的基因结构及表型 |
| 临床生物化学 |
|
|
|
各种生物都能通过生殖产生子代,子代和亲代之间,不论在形态构造或生理功能的特点上都很相似,这种现象称为遗传(heredity)。但是,亲代和子代之间,子代的各个体之间不会完全相同,总会有所差异,这种现象叫变异(variation)。遗传和变异是生命的特征。遗传和变异的现象是多样而复杂的,正因为如此,才导致生物界的多种多样性,生物体所具有的遗传性状称为表型或表现型(phernotype)。生物体所具有的特异基因成分称为基因型(genotype)。表型是基因型与环境因素相互作用的结果。遗传物质是相对稳定的,但是又是可变的,遗传物质的变化以及由其所引起表型的改变,称为突变(mutation)。遗传物质突变包括染色体畸变和基因突变。基因突变是染色体中某一点上发生化学改变,所以又称为点突变(pointmutation)。基因结构和遗传表型的研究是深入了解脂蛋白代谢缺陷症的分子生物学基础,逆向遗传学方法(reversegeneticapproach)则使其有可能在蛋白质水平系统地分析结构和功能的关系。现已采用一个特定的cDNA探针从基因文库中筛选所需要的基因进行cDNA克隆,测定其核苷酸序列,然后从核苷酸序列推断蛋白质氨基酸序列。目前,已分离出许多与动脉粥样硬化有关的脂蛋白的cDNA克隆,并将其蛋白质一级结构的氨基酸排列顺序和基因的核苷酸顺序测出。现已查明,ApoAⅠ、AⅣ、E、B、CⅡ和(a)都存在着异构体,也就是说存在着各种不同的表型或基因型,并可分别从蛋白质水平和核酸水平进行分型。现分别介绍几种主要载脂蛋白的基因结构。
(二)载脂蛋白基因结构特点
人血浆中载脂蛋白的结构及功能,经过近十年的深入研究,已了解得较为清楚。大部分载脂蛋白的基因和cDNA都已得到分离和确定,其核苷酸顺序也进行了测定。除ApoAⅣ,B、(a)外,它们的共同特点是含有三个内含子(intron)和四个外显子(exon),其内含子插入外显子的位置大致相同,基本上按照生理功能的不同,将其加以分隔。第一个内含子把5′-末端的非翻译区和翻译区分开;第二个内含子把信号肽编码(singnalpeptide)和功能蛋白编码区分开;第三个内含子则把原肽编码区和成熟肽编码区分开。这些基因的第一、二、三外显子的核苷酸数量也相差无几,第四个外显子核苷酸数量不同而导致各种载脂蛋白基因长度不同。从生物进化角度考虑,上述载脂蛋白基因结构相似性,提示可能来源于一个共同的祖先,即ApoCⅠ基因。ApoAⅣ与其他载脂蛋白基因结构不同,它只含有三个外显子。载脂蛋白基因结构的另一特点是几个基因相接很近,定位于同一染色体的一个位点上或附近,呈紧密连锁状态。如ApoAⅠ、CⅢ和AⅣ基因位于第11号染色体长臂2区,形成一个约15kb的基因簇。还有一个紧密连锁的基因簇是ApoE、CⅠ和CⅡ基因,同位于第19号染色体长臂3区,见图4-4。
ApoA-Ⅱ基因定位于第1号染色体长臂2区,ApoB基因定位于第2号染色体短臂2区,Apo(a)基因定位于第6号染色体长臂2区。
(三)载脂蛋白基因结构
1.ApoAⅠApoAⅠ基因长1863bp,含有三个内含子,第一个内含子位于5′端非翻译区;第二个内含子位于翻译区的AⅠ前肽区内;第三个内含子插入翻译成熟AⅠ第43氨基酸残基处。ApoAⅠ基因含有四个外显子,分布于ApoAⅠ基因的不同区域,ApoAⅠ基因与ApoCⅢ、AⅣ基因相连成簇,CⅢ基因居中,转录方向与AⅠ和AⅣ基因相反。位于AⅠ和CⅢ基因共同3′区的DNA序列,可能参与对AⅠ基因的转录调控。
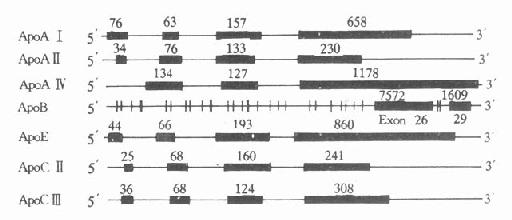
图4-4 人载脂蛋白AⅠ、AⅡ、AⅣ、B100、
CⅡ、CⅢ和E基因结构示意图
粗线代表外显子,粗线之间的细线代表内含子,粗线上缘数字
代表该段核苷酸数目
2.ApoBApoB族位于2号染色体P23→Pter区,是由非翻译区、编码区、TAA终止密码子和一个3′端的非翻译区组成。ApoB100基因全长43kb,含29个外显子和28个内含子见图4-4,其中第26和第29两个外显子特别长,分别含有7552和1905bp,外显子2最短,仅39bp(从211-249)。内含子则以第27个为最短(107bp)。人群中至少有14种不同的3′端高变异等位基因区,75%的人群在此区是不均一的。
ApoB48和ApoB100除了在结构上有关外,ApoB48的形成机制目前尚无完全一致的看法,主要认为有合成ApoB48的基因存在。1987年被发现ApoB48是由ApoB100通过一种新的机制涉及到mRNA的编辑而产生的。在测定从人小肠基因库分离的ApoBcDNA的序列时发现,小肠ApoBcDNA的第6666个核苷酸为T,而从肝分离的ApoBcDNA克隆在此位置为C。将T替换C则6666处产生一终止编码(TAA),TAA替换CAA编码使ApoB100的2153位氨基酸应为Gln,预示血浆中存在的ApoB48应是相当于ApoB100的2153氨基末端为Gln。这一预测后来得到实验证实,并发现核苷酸上6666的替换C→T只发生在小肠的mRNA上,而不发生在小肠基因组(genomic)DNA上,因此这是转录以后的一种特殊形成的编辑小肠mRNA的结果.
3.ApoE人ApoE基因位于19号染色体长臂3区,含有四个外显子和三个内含子。
1975年首先观察到ApoE的多态性,利用等电聚焦电泳和SDS-PAGE可以确认ApoE的多态性。实验表明,ApoE有三种异构体(isoform)即E2、E3和E4。有的人只含有一种主要异构体即纯合子,有的人可含二种主要异构体为杂合子。由此可见,人群中可有六种不同的表现。根据ApoE表型提出ApoE基因模型认为,ApoE的合成是由位于一个基因位点上的三个等位基因所控制,即E2、E3和E4,每一个等位基因对应于一个主要异构体,产生三种纯合子(E2/2,E3/3,E4/4)和三种杂合子(E2/3,E2/4,E3/4)共六种常见表型,另外,还有极少见的异构体。一般认为,次要异构体是由主要异构体翻译后,经唾液酸糖化修饰后转变而来。ApoE3/3型又称野生型。ApoE的基因序列的112位和158位两种氨基酸残基即精氨酸(Arg)和半胱氨酸(Cys)的交换决定了异构体的种类。ApoE4在这两个位置上都是Arg;E2都是Cys;112和158位是Arg者为ApoE3异构体。自然人群中,基因频率(3)分布最高,ApoE3/3表型分布约70%,见图4-5。
4.ApoC族ApoCⅡ基因有3347bp,含有4个外显子和3个内含子。ApoCⅡ的羧基末端氨基酸序列是激活脂蛋白脂肪酶的活性功能区域。ApoCⅢ基因含有3133bp,有4个外显子和3个内含子。

图4-5 人ApoE三种主要异构体的氨基酸残基及基因密码的改变位置
5.Apo(a)运用cDNA探针进行染色体定位研究时发现,Apo(a)的基因位点在人第6号染色体长臂2区6-7带间,与血纤溶酶原(PLG)的基因位点有部分重叠。测定PLG基因跨距为525kb,由18个内含子与19个外显子组成,5个Kringle结构由各自两个外显子编码。Apo(a)cDNA分析表明,Apo(a)与PLG的基因有很多相似之处。
通过家系研究,目前已发现Apo(a)基因位点中至少有26个等位基因与多态性有关。这些等位基因至少表达有34种Apo(a)异构体。
| 关于“临床生物化学/载脂蛋白的基因结构及表型”的留言: | |
|
目前暂无留言 | |
| 添加留言 | |