临床生物化学/LDL受体
| 医学电子书 >> 《临床生物化学》 >> 血浆蛋白及其代谢紊乱 >> 脂蛋白受体 >> LDL受体 |
| 临床生物化学 |
|
|
最先从牛肾上腺分离出LDL受体,以后又分离了编码牛LDL受体羟基末端1/3氨基酸的cDNA,并初步阐明了牛LDL受体的cDNA,并且推导出人LDL受体的氨基酸序列。
(一)LDL受体结构
LDL受体是一种多功能蛋白,由836个氨基酸残基组成36面体结构蛋白,分子量约115ku,由五种不同的区域构成,各区域有其独特的功能,见图4-6。
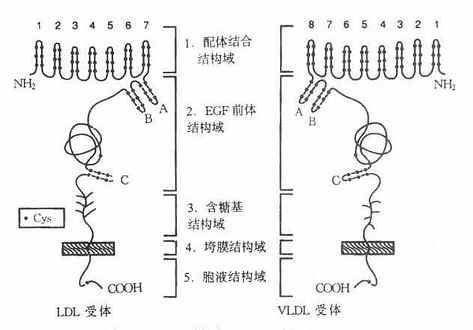
图4-6 LDL受体与VLDL受体结构示意图
1.配体结合结构域配体结合结构域由292个氨基酸残基组成,其中共有47个半胱氨酸(Cys)。含有七个由40个残基组成的与补体Cb和Cq类似的重复序列,每个重复系列中有6个半胱氨酸残基,所有42个半胱氨酸残基均已构成二硫键,重复序列2,3,6,7是结合LDL所必需,其中任何一种发生突变,均使受体丧失结合LDL的能力。重复序列5则与结合β-VLDL有关,若该序列突变时,受体结合β-VLDL的能力丧失60%。该受体不仅能结合LDL,还能结合VLDL、β-VLDL和VLDL残粒,它不仅能识别ApoB100,也可识别含ApoE的脂蛋白。ApoE、B100为LDL受体的配体,因此,LDL受体又称为ApoB100E受体。
2.EGF前体结构域该域约由400个氨基酸残基组成的肽段,有五个重复序列,每个重复序列包括25个氨基酸残基。EGF前体结构域与小鼠上皮细胞生长因子(epidermalgrowthfactor,EGF)前体有同源性,这一区域因此而得名。体外实验证实,这个区域的肽段,属于细胞膜外结构蛋白,起着支撑作用。
3.糖基结构域由58个氨基酸残基组成,是紧靠细胞膜面的肽段,有18个丝氨酸或苏氨酸,构成O-连接糖链,对LDL受体也有支撑作用。
4.跨膜结构由22个氨基酸残基组成,富含疏水氨基酸残基,属于跨膜蛋白,起着固系于细胞膜中的“抛锚”作用。这个区域若有缺陷则影响受体的细胞外分泌。
5.胞液结构域位于细胞膜的胞质侧,由50个氨基酸残基组成,C-末端位于胞质并“深埋”于胞质之中。
(二)LDL受体基因结构及功能
1.受体亲和性含ApoB100的脂蛋白可以与LDL受体以高亲和力结合,肠道分泌的ApoB48不是LDL受体的配体,所以肝脏不能清除完整的CM。
2.基因结构人LDL受体基因长度45ku,由18个外显子和17个内含子组成。
3.LDL受体途径LDL受体广泛分布于肝、动脉壁平滑肌细胞、肾上腺皮质细胞、血管内皮细胞、淋巴细胞、单核细胞和巨噬细胞,各组织或细胞分布的LDL受体活性差别很大。
LDL或其他含ApoB100、E的脂蛋白如VLDL、β-VLDL均可与LDL受体结合,内吞入细胞使其获得脂类,主要是胆固醇,这种代谢过程称为LDL受体途径(LDLreceptorpathway)。该途径依赖于LDL受体介导的细胞膜吞饮作用完成,如图4-7所示。当血浆中LDL与细胞膜上有被区域(coatedregion)的LDL受体结合(第1步),使其出现有被小窝(coatedpit)(第2步),并从膜上分离形成有被小泡(coatedvesicles)(第3步),其上的网格蛋白(clathrin)解聚脱落,再结合到膜上(第4步),其内的pH值降低,使受体与LDL解离(第5步),LDL受体重新回到膜上进行下一次循环(第6、7步)。有被小泡与溶酶体融合后,LDL经溶酶作用,胆固醇酯水解成游离胆固醇和脂肪酸,甘油三酯水解成脂肪酸,载脂蛋白B100水解成氨基酸。LDL被溶酶体水解形成的游离胆固醇再进入胞质的代谢库,供细胞膜等膜结构利用。胞内游离胆固醇在调节细胞胆固醇代谢上具有重要作用;若胞内浓度升高,可能出现下述种情况:①抑制HMGCoA还原酶,以减少自身的胆固醇合成;②抑制LDL受体基因的表达,减少LDL受体的合成,从而减少LDL的摄取,这种LDL受体减少的调节过程称为下调(downregulation);③激活内质网脂酰基CoA胆固醇酰转移酶(Acyl-CoAcholesterolacyltransferase,ACAT),使游离胆固醇在胞质内酯化成胆固醇酯贮存,以供细胞的需要。经上述三方面的变化,用以控制细胞内胆固醇含量处于正常动态平衡状态。血浆中胆固醇主要存在于LDL中,而65%-70%的LDL是依赖肝细胞的LDL受体清除。肝的LDL受体还影响LDL的合成速率及VLDL代谢。曾经认为人VLDL几乎全部在血循环中转变为LDL,LDL再被肝外组织摄取。现在经大鼠和兔实验研究表明,仅有15%以下转变为LDL,人则是小于50%的VLDL转变为LDL,大部分VLDL是以VLDL或VLDL残粒的形成被肝摄取。VLDL残粒与肝受体的亲和力比VLDL大很多。所以VLDL残粒被肝清除的速率比VLDL快。VLDL残粒大部分被肝清除,一小部分在肝脂酶作用下水解除去甘油三酯而转变成LDL。LDL受体还在乳糜微粒代谢中起一定作用。乳糜微粒中的ApoB48不能识别ApoB100E受体,所以肝不能清除完整的乳糜微粒。CM中虽有少量ApoE,因含有丰富的ApoC,可掩盖ApoE,而阻碍其与肝的ApoB、E受体结合,血液中乳糜微粒被脂蛋白脂肪酶水解去除其大部分甘油三酯核心后,同时丧失部分ApoC、A,生成乳糜微粒残粒后除去了阻碍ApoE与受体结合的因素,其残粒可迅速被肝清除,约有一半是通过LDL受体,另一半通过LDL受体相关蛋白代谢,其半寿期短。
总之,LDL受体主要功能是通过摄取Ch进入细胞内,用于细胞增殖和固醇类激素及胆汁酸盐的合成等。
| 关于“临床生物化学/LDL受体”的留言: | |
|
目前暂无留言 | |
| 添加留言 | |