生理学/自律细胞的跨膜电位及其形成机制
| 医学电子书 >> 《生理学》 >> 血液循环 >> 心肌的生物电现象和生理特征 >> 心肌细胞的生物电现象 >> 自律细胞的跨膜电位及其形成机制 |
| 生理学 |
|
|
|
在没有外来刺激时,工作细胞不能产生动作电位,在外来刺激作用下,产生一次动作电位,但两次动作电位之间膜电位是稳定不变的。而在自律细胞,当动作电位3期复极未期达到最大值(称最大复极电位)之后,4期的膜电位并不稳定于这一水平,而是立即开始自动除极,除极达阈电位后引起兴奋,出现另一个动作电位。这种现象,周而复始,动作电位就不断地产生。出现于4期的这种自动除极过程,具有随时间而递增的特点,其除极速度远较0期除极缓慢;不同类型的自律细胞4期除极速度参差不一,但同类自律细胞4期除极速度比较恒定。这种4期自动除极(亦称4期缓慢除极或缓慢舒张期除极),是自律细胞产生自动节律性兴奋的基础。
根据细胞膜除极的跨膜电流的基本规律可分析自律细胞4期自动除极形成的机制。不难推测,自律细胞由于净外向电流使膜复极(3期)达最大复极电位后,在4期中又出现一种逐渐增强的净内向电流,从而使膜内正电位逐渐增加,膜便逐渐除极。这种进行性净内向电流的产生,有以下三种可能的原因:①内向电流的逐渐增强;②外向电流的逐渐衰退;③两者兼有。不同类型的自律细胞,4期自动除极都是由这种进行性净内向电流所引起,但构成净内向电流的离子流的方向和离子本质并不完全相同。
1.浦肯野细胞浦肯野细胞是一种快反应自律细胞。作为一种快反应型细胞,它的动作电位的形态与心室肌细胞相似,产生的离子基础也基本相同。
关于浦肯野细胞4期自动除极形成的机制,80年代研究资料表明,在浦肯野细胞,随着复极的进行,导致膜复极的外向K+电流逐渐衰减,而同时在膜电位4期可记录到一种随时间推移而逐渐增强的内向电流(If)(图4-7)。If通道在动作电位3期复极电位达-60mV左右开始被激活开放,其激活程度随着复极的进行、膜内负电性的增加而增加,至-100mV左右就充分激活。因此,内向电流表现出时间依从性增强,膜的除极程度因而也随时间而增加,一旦达到阈电位水平,便又产生另一次动作电位,与此同时,这种内向电流在膜除极达-50mV左右因通道失活而中止。可见,动作电位的复极期膜电位本身是引起这种内向电流启动和发展的因素,内向电流的产生和增强导致膜的进行性除极,而膜的除极一方面引起另一次动作电位,一方面又反过来中止这种内向电流。这一连串的过程是自律细胞“自我”启动、“自我”发展,又“自我”限制的,由此可以理解为什么自律细胞能够自动地、不断地产生节律性兴奋。
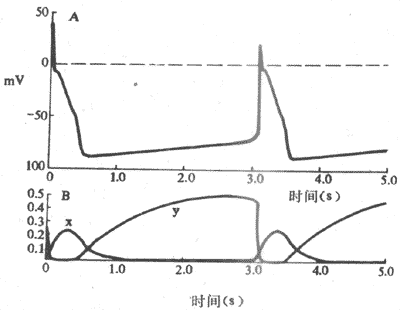
图4-7 浦肯野细胞起搏机制
A;跨膜电位 B:由х闸门控制的Ik衰减以及由у闸门控制的If,两者在形成起搏电位中的相对关系
这种4期内向电流,通常称为起搏电流,其主要离子成分为Na+ ,但也有K+参与。由于使它充分激活的膜电位为-100mV,因而认为,构成起搏内向电流的是一种被膜的超极化激活的非特异性内向(主要是是Na+)离子流,标志符号为If。If的通道允许Na+通过,但不同于快Na+通道,两者激活的电压水平不同;If可被铯(Cs)所阻断,而河豚毒却不能阻断它。目前,关于If及其通道的研究资料尚有若干不能充分予以解释的疑点,对If的进一步研究正受到心肌电生理学者们的高度关注。
2.窦房结细胞的跨膜电位及其形成机制 窦房结含有丰富的自律细胞,动作电位复极后出现明显的4期自动除极,但它是一种慢反应自律细胞,其跨膜电位具有许多不同于心室肌快反应细胞和浦肯野快反应自律细胞的特征:①窦房结细胞的最大复极电位(-70mV)和阈电位(-40mV)均高于(电位较正)浦肯野细胞;②0期除极结束时,膜内电位为0mV左右,不出现明显的极化倒转;③其除极幅度(70mV)小于浦肯野细胞(为120mV),而0期除极时程(7ms左右)却又比后者(1-2ms)长得多。原因是窦房结细胞0期除极速度(约10V/s)明显慢于浦肯野细胞(200-1000V/s),因此,动作电位升支远不如后者那么陡峭;④没有明显的复极1期和平台期;⑤4期自动除极速度(约0.1V/s)却比浦肯野细胞(约0.02V/s)要快,记录曲线上窦房结细胞4期膜电位变化的斜率大于浦肯野细胞.图4-8显示心室肌快反应细胞与窦房结细胞跨膜电位变化的差别。
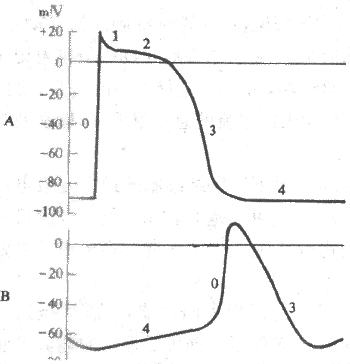
图4-8 心室肌(A)与窦房结(B)细胞跨膜电位的比较
窦房结细胞的直径很小,进行电生理研究有一定困难。直到70年代中期,才开始在窦房结小标本上采用电压钳技术对其跨膜离子流进行了定量研究,但目前尚未能充分阐明它的跨膜电位,尤其是4期起搏电流的离子基础。学者们观察到,窦房结细胞0期除极不受细胞外Na+浓度的影响,对河豚毒很不敏感;相反,它受细胞外Ca2+浓度的明显影响,并可被抑制钙通道的药物和离子(如异搏定、D-600和Mn2+等)所阻断。据此可以认为,引起窦房结细胞动作电位0期除极的内向电流是由Ca2+负载的。这种内向电流被称为第二内向电流;而引起快反应细胞(心室肌、心房肌和浦肯野细胞)0期除极的快Na+内流称为第一内向电流。根据已有的研究资料,可将窦房结细胞动作电位的形成过程描述如下:当膜电位由最大复极电位自动除极达阈电位水平时,激活膜上钙通道,引起Ca2+内向流(Ica),导致0期除极;随后,钙通道逐渐失活,Ca2+内流相应减少;另一方面,在复极初期,有一种K+通道被激活,出现K+外向流(Ik)。Ca2+内流的逐渐减少和K+外流的逐渐增加,膜便逐渐复极。由“慢”通道所控制、由Ca2+内流所引起的缓慢0期除极,是窦房结细胞动作电位的主要特征,因此,相应称为慢反应细胞和慢反应电位,以区别于前述心室肌等快反应细胞和快反应电位。
窦房结细胞的4期自动除极也由随时间而增长的净内向电流所引起,但其构成成分比较复杂,是几种跨膜离子流的混合。目前已知,在窦房结细胞4期可以记录到三种膜电流,包括一种外电流和两种内向电流,不过它们在窦房结细胞起搏活动中所起作用的大小以及起作用的时间有所不同。
(1)Ik通道的激活和逐渐增强所造成的K+外向流,是导致窦房结细胞复极的原因。Ik通道在膜复极达-40mV时便开始逐渐失活,K+外流因此渐渐减少,导致膜内正电荷逐渐增加而形成4期除极。目前认为,由于Ik通道的时间依从性逐渐失活所造成的K+外流进行性衰减,是窦房结细胞4期自动除极的最重要的离子基础(图4-9);
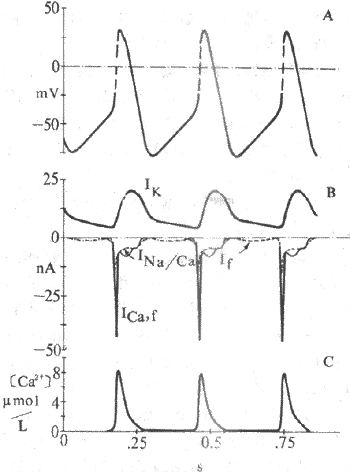
图4-9 窦房结动作电位和起搏电位的离子机制
A.跨膜电位 B.越膜电位C. 胞浆Ca2+浓度表示动作电位升支由Ica,
f构成,起搏电位 由Ik和If及Ina/Ca构成
(2)If:If是一种进行性增强的内向离子(主要为Na+)流,在浦肯野细胞起搏活动中,If起着极重要的作用,而Ik衰减的作用很小。与此恰相反,窦房结细胞4期虽也可记录到If ,但它对起搏活动所起的作用不如Ik衰减。实验证明,用Cs2+选择性阻断If后,窦房结自发放频率仅轻度减少;对家兔窦房结细胞4期净内向电流的总幅值而言,Ik衰减与If两者所起作用的比例为6:1。
If通道的最大激活电位为-100mV左右,而正常情况下窦房结细胞的最大复极电位为-70mV,在这种电位水平下,If通道的激活十分缓慢,这可能是If在窦房结4期除极过程中所起作用不大的原因。若窦房结细胞发生超级化时,If则可能成为起搏电流中的主要成分。
(3)此外,窦房结细胞4期中还存在一种非特异性的缓慢内向电流,在膜除极达-60mV时被激活,可见,它在自动除极过程的后1/3期间才起作用。这种缓慢内向电流可能是生电性Na+-Ca2+交换的结果(Na+-Ca2+交换时,心肌细胞排出一个Ca2+,摄入3个Na+,出/入细胞正电荷之比为2:3,形成内向电流)。
心肌细胞的类型 除了按照功能和电生理特性将心肌细胞分为工作细胞和自律细胞之外,还可以根据其生物活动的特征,特别是动作电位0期除极的速度,将心肌细胞分为快反应细胞和慢反应细胞,其动作电位相应称为快反应电位和慢反应电位;然后再结合其自律性,可将心肌细胞分为以下四种类型:
快反应非自律细胞:包括心房肌细胞和心室肌细胞;
快反应自律细胞:浦肯野自律细胞;
慢反应自律细胞:窦房结自律细胞,以及房结区和结希区的自律细胞;
慢反应非自律细胞:结区细胞。
心肌细胞的跨膜离子流 由多种离子通过细胞膜上的特异性或非特异性离子通道以及经离子交换转运形成的跨膜离子运动,是心肌细胞跨膜电位形成的基础。心肌细胞跨膜离子流的种类众多,性质很复杂,近年又取得许多进展。由于在实验研究中对这些离子通道的命名和认识比较混乱,甚至前后矛盾,故将目前比较肯定的主要几种离子流和通道的名称、存在的部位和作用归纳介绍如下:
1.内向离子流
(1)INa:称为快速Na+流,存在于快反应细胞,是引起快反应细胞0期除极的内向电流(称第一内向电流)的离子基础。
(2)Isi:称为缓慢内向电流,也称第二内向电流,过去认为它是一种由慢通道控制的缓慢Ca2+流,故标志为ICa,其功能是构成快反应细胞的平台期和慢反应细胞的除极期。新近的研究结果在很大程度上修正了对Ica的认识。目前认为,第二内向电流并不是单一的Ca2+流,而是由特性各异的三个组分所构成;第一组分称Ica.f,二、三组分别称Is1.2和Ica.s。其中①Ica.f是一种快速Ca2+流,其通道激活和失活的速度远比已往所认为的要快得多。它融合于快反应细胞Na+内流的最后部分,共同形成动作电位升支的上段,而对平台期的作用很小。它是慢反应细胞除极的离子基础。对工作细胞而言,肌浆网Ca2+再生性释放帅细胞外Ca2+进入胞浆触发的,既然Ica.f的激活十分迅速,那么Ca2+释放的触发有及兴奋-收缩耦联的启动速度都比以往认为的要快得多;②第三组分Ica.s,是一种较Ica.f微弱而缓慢的Ca2+流,主要作用是维持快反应细胞平台期;③Is1.2称第二内向电流第二组分,其离子本质不甚清楚,可能是Na+-Ca2+交换的生电电流,故也称为INa.Ca,在平台期起作用;慢反应自律细胞4期自动除极晚期也有它的作用。
(3)If:超极化激活的非特异性内向离子流,主要由Na+携带,存在于自律细胞4期。If是决定浦肯野快反应自律起搏活动的主要负离子流,而在窦房结慢反应自律细胞起搏活动中,其作用不如Ik衰减。
2.外向离子流
(1)Ikl:存在于快反应细胞,是决定快反应工作细胞静息电位的离子流,并在复极2期和3期起复极作用;
(2)Ik:这种外向电流主要由K+携带,但也有Na2+参加,不是单纯的K+流,故又称Ix,在快反应细胞复极3期起重要作用,故又称为复极电流,因而也决定着浦肯野细胞的最大复极电位;Ix也存在于慢反应自律细胞,促使膜复极,4期内呈进行性衰减,是形成4期除极的主要离子基础;
(3)Ito:是快反应细胞1期复极的离子流,离子成分主要为K+,也有Na+参与。
3.化学门控离子通道 近年相继证实,在心肌细胞膜上,除了电压门控通道外,还存在化学门控离子通道;它们的作用并不在于维持和产生正常跨膜电位(静息电位和动作电位),但对于心肌细胞活动的调节以及异常电活动的电生产有着特别重要的意义。重要的化学门控离子通道有以下三种;
(1)乙酰胆碱控制的K+通道(Ik-ACh):早年曾认为ACh激活的是Ikl,近年发现是通过G蛋白激活开放了另一种与在生物物理学特性和生理学特性上均不相同的K+通道。
(2)ATP依从性K+通道(Ik-ATP):ATP的作用并不是分解供能激活此通道,而是维持此通道在正常情况下处于关闭状态。当心肌细胞内ATP降到临界水平以下时(如心肌缺血时),此种特殊的K+通道开放。大量K+外漏以致缺血心肌细胞局部高钾而引起除极,诱发心律失常。硫脲类药物可阻断此通道。
(3)INa–k—Ca2+通道:是细胞内Ca2+增高时激活的一种非特异性正离子通道,载流离子是Na+和K+,形成一过性内向离子流(Iti)。实验表明,在某些情况下,浦肯野细胞在动作电位复极后可产生一种除极电位(延迟后除极电位),当它达阈电位时就可以诱发另一个新的动作电位,形成异位搏动。Iti就是延迟后除极电位的离子基础。洋地黄中毒,细胞外低K+或低Na+,以及咖啡因、儿茶酚胺等可引起细胞内Ca2+超负荷的因素,均可诱发或加强Iti和延迟后除极电位。
| 关于“生理学/自律细胞的跨膜电位及其形成机制”的留言: | |
|
目前暂无留言 | |
| 添加留言 | |