医学免疫学/免疫球蛋白基因的结构和抗体多样性
| 医学电子书 >> 《医学免疫学》 >> 免疫球蛋白分子 >> 免疫球蛋白基因的结构和抗体多样性 |
| 医学免疫学 |
|
|
|
Ig分子是由三个不连锁的Igκ、Igλ和IgH基因所编码。Igκ、Igλ和IgH基因定位于不同的染色体上(表2-5)。编码一条Ig多肽链的基因是由在胚系中多个分隔的DNA片段(基因片段)经重排而形成的。1965年Dreyer和Bennet首先提出假说,认为Ig的V区和C区是由分隔存在的基因所编码,在淋巴细胞发育过程中这两个基因发生易位而重排在一起。1976年日本学者利根川进应用DNA重组技术证实了这一假说。利根川进由此获得1987年医学和生理学诺贝尔奖。
表2-5 免疫球蛋白基因定位
| 编码多肽链 | 基因符号(人) | 基因染色体定位 | ||
| 人 | 小鼠 | |||
| κ轻链 | Igκ | 2 | 6 | |
| λ轻链 | Igλ | 22 | 16 | |
| 重链 | IgH | 14 | 12 | |
一、Ig重链基因的结构和重排
(一)重键V区基因
H链V区是由V、D、J三种基因片段经重排后组成。
1.H链V区基因组成
(1)V基因片段:小鼠VH基因段约为250~1000,人的VH基因片段约为100。V基因片段编码VH的信号序列和V区靠N端98个氨基酸残基,包括CDR1和CDR2。
(2)D基因片段:D是指多样性(diversity)。D基因片段仅存在于H链,不存在于L链。小鼠DH共有12个片段,人的DH片段的数目还不完全清楚,可能有10~20个左右。D片段编码H链CDR3中大部分氨基酸残基。
(3)J基因片段:J是连接(joining)的意思。JH连接V基因片段和C基因片段。小鼠JH有4个,人有9个JH片段,其中6个是有功能的。J基因片段编码CDR3的其余部分氨基酸残基和第4个骨架区。
2.H链V区基因的移位 首先发生D与J基因片段的连接形成D-J,然后V基因片段与D-J基因片段连接。H链V区基因的易位和连接是通过七聚体-间隔序列-九聚体识别信号和重组酶而完成的。
(二)重链C区基因
1.C基因片段小鼠H链区基因片从5’端到3’排列的顺序是Cμ-Cδ-Cγ3-Cγ1-Cγ2b-Cε-Cα2,人H链C区基因的顺序为Cμ-Cδ-Cγ3-Cγ1-Cε2(pseudo基因)- Cα2- Cγ2-Cγ- Cε- Cα2(图2-13,14)。
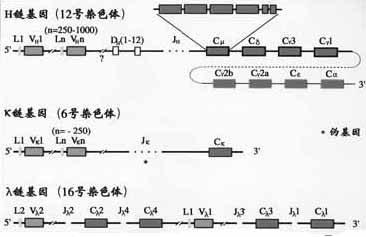
图2-13小鼠Ig基因结构
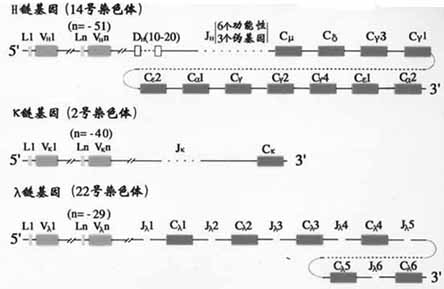
图2-14 人Ig基因结构
2.Ig类别转换(class switch) 是指一个B细胞克隆在分化过程中,V基因不变,而CH基因片段不同重排,比较CH基因片段重排后基因编码的产物,其V区相同,而C区不同,即识别抗原的特异性相同,而Ig的类或亚类发生改变。Ig可能是通过缺失模式(deletion model)和RNA剪接(splicing)两种机制来实现类别的转换。
(三)膜表面Ig重链基因
膜表面Ig(Sm Ig)是B细胞识别抗原的受体。(Sm Ig)和分泌性Ig的H链结构相类似,所不同的是smIgH名字的羧基端多含一段穿膜的疏水性氨基酸残基和胞浆区。因此SmIgH链的转录本(transcript)要比分泌性IgH链转录本多1~2个外显子。编码H链的羧基端部分,其氨基酸残基的的数目视H链不同而有差异,如在小鼠或人SmIgμ链的这一部分长约41个氨基酸残基,而小鼠SmIgε链此区域却有72个氨基酸残基。这个区域包括三个部分:①一个酸性间隔子,与H链最后一个CH功能区相同,位于胞膜外侧;②含26个氨基酸残基的疏水区,为穿膜部分;③胞浆内部分,3~28个氨基酸残基不等。
二、Ig轻链基因的结构和重排
在IgH链基因重排后,L链可变区基因片段随之发生重排。在L链中,κ链基因先发生重排,如果κ基因重排无效,随即发生λ基因的重排。L链匠CDR1、CDR2和大部分CDR3由Vκ或Vλ基因片段所编码(Vκ编码95个氨基酸残基),Jκ或Jλ基因片段编码CDR3的其余部分和第四个骨架区(Jλ编码从96位到108位氨基酸)。L链无D基因片段。
(一)κ链基因的结构和重排
κ链基因是V基因片段(Vκ)、J基因片段(Jκ)和C基因片段(Cκ)重排后组成。小鼠Vκ基因片段约有250,Jκ有5个(其中4个功能),Cκ只有1个。人Vκ基因片段约有100个,Jκ有5个。Cκ也只有1个。Vκ与Cκ之间以随机方式发生重排。
(二)λ链基因的结构和重排
κ链基因也是由Vλ、Jλ和Tλ基因片段经重排后组成。小鼠Vλ基因片段有3个:Vλ1、VλX;4个Jλ和4个Cλ基因片段,分为(Jλ2Cλ2,Jλ4Cλ和Jλ3Cλ3,Jλ1Cλ1)两组。它们的基因重排比较复杂。人Vλ约有100个,至少有6个Cλ与各自的J基因片段相连,人λ链确切的重排情况还不清楚。
三、抗体多样性的遗传学基础
机体对外界环境中种类众多抗原刺激可产生相应的特异性抗体,推算出抗体的多样性在107以上。抗体多样性主要由基因控制。
1.胚系(germ line)中众多的V、D、J基因片段 在胚系上,尚未重排的Ig基因片段数量相当多,这是生物在长期进化中形成的。表2-6例举了小鼠H链和L链重排的多样性以及H链和κ链相互随机配对所推算的多样性数目。
表2-6 小鼠Ig多样性(举例)
| 多肽链 | 基因片段数 | V区基因重组方式 | 经重排的随机配对后* 推算的多样性数目 |
|||
| V | J | |||||
| H链 | 1000 | 12 | 4 | V-D-J | 4.8×104 | 4.8×107 |
| κ链 | 250 | - | 4 | V-J | 1.0×103 | |
- 多样性数目不包括VDJ连接多样性、N区插入和体细胞突变所增加的多样性数目
2.VDJ连接的多样性在L链基因重排过程中V-J连接位点有一定的变异范围,例如VL基因片段3'端5个核苷酸CCTCC和JL基因片段5'端4个核苷酸GTGG连接时,总共9个核苷酸中只有6个核苷酸编码L链第95、96位氨基酸,因此可产生8种不同的连接方式。在H链基因重排过程中K-J以及V-D-J连接时都可有连接多样性的存在。
3.体细胞突变(somaticmutation)体细胞在发育过程中可发生基因突变。以长期体外培养的B细胞前体为例,每个细胞每个碱基对的突变率约为1~43×10-5,这种点突变主要发生在V基因。体细胞突变扩展了原有胚系众多基因片段重排的多样性。
4.N区的插入在IgH链基因片段重排过程中,有时可通过无模板指导的机制(nontempletdirected mechanism),在重组后D基因片段的两侧即VH-DH或DH-JH连接处额外插入称为N区的几个核苷酸。N区不是由胚系基因所编码。在N区插入前,先通过外切酶切除VH-DH或DH-JH连接处几个碱基对,然后通过末端脱氧核苷酸转移酶(terminal deoxynucleotidyl transferase,TdT)连接上N区。由于额外插入了N区,可发生移码突变(fuame shift mutation),使插入部位以及下游的密码子发生改变,从而编码不同的氨基酸,大大地增加了抗体的多样性。
5.L链H链相互随机配对 如表2-6所示,小鼠H链和κ链随机配对后推算其多样性可达4.8×107,如果再加上H链与λ链的随机配对其多样性应更多了。
| 关于“医学免疫学/免疫球蛋白基因的结构和抗体多样性”的留言: | |
|
目前暂无留言 | |
| 添加留言 | |